
Problèmes résolus de génétique
"Ne me dites pas que ce problème est difficile. S'il n'était pas difficile, ce ne serait pas un problème." Maréchal FOCH, 1851-1929
Monobridisme
Il existe différents types de radis: ronds, longs ou ovales. Des croisements entre plantes à radis long et plantes à radis ovale ont produit 159 plantes à radis long et 156 plantes à radis ovales. D'autres croisements entre plantes à radis rond et plantes à radis ovale ont produit 199 plantes à radis rond et 203 plantes à radis ovales. Deux autres types de croisements ont été pratiqués: l'un entre plantes à radis long et plantes à radis rond a fourni 576 plantes à radis ovale; l'autre entre plantes à radis ovale a donné 121 plantes à radis long, 243 plantes à radis ovale et 119 plantes à radis rond.
Déduisez-en le mode de transmission du caractère de la forme du radis.
Il existe différents types de radis: ronds, longs ou ovales. Ces phénotypes correspondent soit à l'expression de génotypes homozygotes (il existerait dès lors pour le caractère "forme du radis", les allèles "rond", "long" ou "ovale"), soit à l'expression de génotypes hétérozygotes, fait de deux allèles différents. La seule lecture de la dernière information de l'énoncé ("le croisement entre plantes à radis ovale a donné 121 plantes à radis long, 243 plantes à radis ovale et 119 plantes à radis rond") montre les proportions typiques (1/4, 2/4 et 1/4) des phénotypes obtenus en première génération à l'issue du croisement entre deux lignées pures distinctes par un caractère, soit l'expression de la première loi de Mendel dans le cas de deux allèles codominants.
Les deux allèles codominants du caractère "forme du radis" sont donc "rond" et "long", le phénotype "ovale" étant l'expression du génotype hétérozygote.
La lecture des autres informations de l'énoncé ne fait que confirmer la réponse.
***
La fibrose kystique est une maladie autosomique récessive caractérisée surtout par une insuffisance pancréatique et une accumulation de mucus dans les voies respiratoires.
a) Déterminez, d'après la figure précédente, si la fibrose kystique dépend d'une gène autosomique (= liée à un chromosome non sexuel) dominant ou récessif (noir= phénotype de fibrose, hommes = carrés, femmes = ronds).
La fibrose kystique dépend d'un allèle récessif. En effet, l'arbre généalogique montre, par deux reprises, des enfants malades de parents sains. Les parents n'exprimant pas la maladie portent l'allèle de celle-ci de façon cachée, c'est-à-dire sous la forme d'un allèle récessif, qu'ils transmettent chacun à leur enfant de génotype homozygote, qui dès lors exprime la maladie.
b) Indiquez le génotype des personnes suivantes:
Femme de la première génération, 1è individu en partant de la gauche:Kk. L'allèle K est la forme normale du caractère, non malade, et k l'allèle récessif déterminant la maladie. Cette femme est normale: elle possède donc au moins un allèle K. Sa fille est malade, de génotype homozygote récessif; cette dernière a donc reçu de sa mère un allèle k.
Homme de la deuxième génération, 4è individu en partant de la gauche: Kk. Cet homme est normal: il possède donc au moins un allèle K. Son fils est malade, de génotype homozygote récessif; ce dernier a donc reçu de son père un allèle k.
Femme de la troisième génération, 1è individu en partant de la gauche: Kk. Cette femme est normale et possède donc au moins un allèle K. Mais sa mère malade et portant deux allèles k récessif lui en a nécessairement transmis un.
c) Comment se fait-il que deux parents normaux aient un enfant souffrant de fibrose kystique? Deux parents normaux ayant un enfant malade lui transmettent chacun un allèle récessif malade ne s'exprimant pas chez les parents sains, mais bien chez l'enfant homozygote kk.
d) Quelle est la probabilité en pourcentage pour ces deux parents d'avoir un enfant souffrant de cette maladie? 1/4. Chaque parent de génotype Kk donne l'allèle k à son enfant selon une (mal)chance sur deux. L'enfant ayant deux parents, la probabilité qu'il hérite de 2 k est donc de 1/2.1/2=1/4
***
Chez les coqs et les poules de basse-cour, la présence d'une crête est due à C, l'allèle c correspondant à l'absence de cet appendice. Un individu à plumes noires est de génotype R?, un individu à plumes rouges étant rr. Un individu noir sans crête de souche pure est croisé avec un individu rouge à crête de souche pure. Les descendants F1 sont croisés entre eux. A la génération F2, les individus noirs à crête sont analysés par testcross. Faites l'étude de ces testcross.
Les F1 d'un croisement RR,cc X rr,CC sont tous Rr,Cc.
Les F2 d'un croisement Rr,Cc x Rr,Cc donnent de nombreux génotypes dont 1/16 de RR,CC, 2/16 de RR,Cc, 2/16 de Rr,CC et 4/16 de Rr,Cc. On peut donc considérer que, parmi les seuls phénotypes noirs à crête de la F2, on rencontre 1/9 de RR,CC, 2/9 de RR,Cc, 2/9 de Rr,CC et 4/9 de Rr,Cc.
Ces quatre génotypes sont soumis à un testcross, soit croisés avec des individus double homozygotes récessifs. A l'issue de ces testcross, les divers phénotypes des enfants apparaissent avec les mêmes proportions que les différents allèles dans les génotypes des individus testés (les différents F2 noirs à crête). On obtient donc dans 1/9 des cas 100% de noirs à crête, dans 2/9 des cas 50% de noirs à crête et 50% de noirs sans crête, dans 2/9 des cas 50% de noirs à crête et 50% de rouges à crête, et dans 4/9 des cas 25% de noirs à crête, 25% de noir sans crête, 25% de rouges à crête et 25% de rouges sans crête.
Finalement, les phénotypes de ces quatre types de testcross apparaissent avec les proportions suivantes:
Noirs à crête: 1.1/9 + 1/2.2/9 + 1/2.2/9 + 1/4.4/9 = 4/9
Noirs sans crête: 1/2.2/9 + 1/4.4/9 = 2/9
Rouges à crête: 1/2.2/9 + 1/4.4/9 = 2/9
Rouges sans crête: 1/4.4/9 = 1/9
On n'a oublié aucun cas, la somme donnant bien 9/9 = 100%
***
L'arbre généalogique ci-dessous montre la transmission des groupes sanguins du système ABo dans une famille:

a) Les sujets A sont-ils de génotype AA ou Ao? Les sujets B sont-ils de génotype BB ou Bo? Est-il toujours possible de répondre à ces questions? Si non, pour quels individus le génotype n'est-il pas assurément déterminé?
b) L'un des sujets est né d'une union illégitime: lequel? Justifiez votre choix.
a) Les sujets A sont tous assurément Ao et les sujets B sont tous assurément Bo sauf l'homme du 4è couple à partir de la gauche de la 2è génération et son garçon A, qui peuvent être AA ou Ao, et sauf la femme d'extrême droite de la 2è génération qui peut être BB ou Bo.
b) Le garçon AB du 4è couple à partir de la gauche de la 2è génération n'est pas né d'un père A, qui n'a pu lui transmettre un allèle B.
***
Dihybridisme
On croise deux variétés d'hibiscus de race pure, différant par plusieurs caractères. Les hybrides F1 croisés entre eux fournissent en F2 la descendance suivante: 82 plantes à corolle ouverte rouge, 28 plantes à corolle fermée rouge, 165 plantes à corolle ouverte rose, 53 plantes à corolle fermée rose, 81 plantes à corolle ouverte blanche et 26 plantes à corolle fermée blanche.
a) Citez les différents allèles en précisant chaque fois s'ils sont dominants ou récessifs.
b) Quels étaient les génotypes et phénotypes des parents P de race pure?
c) Quels étaient les génotype et phénotype des hybrides F1?
d) Quels sont les génotypes des plantes à corolle ouverte rose F2?
e) On désire connaître le génotype de chacune des 165 plantes F2 à corolle ouverte rose. Que faut-il faire? Quel sera le résultat?
a) Pour déterminer les allèles, et la dominance et la récessivité ou la codominance des différents allèles pour chaque caractère, il convient, dans un premier temps, de ne considérer qu'un caractère à la fois.
Considérant le caractère "couleur de la fleur", on observe, par somme des données de l'énoncé, 110 plantes à corolle rouge, 218 plantes à corolle rose et 107 plantes à corolle blanche, soit sensiblement une proportion 1/4 - 2/4 - 1/4. Ces proportions phénotypiques sont caractéristiques des résultats d'un croisement de deux parents de lignée pure différant par un caractère à deux allèles codominants. Pour ce premier caractère, les deux allèles codominants sont donc R = rouge et B = blanc.
Considérant le caractère "ouverture de la fleur", on observe, par somme des données de l'énoncé, 328 plantes à corolle ouverte et 107 plantes à corolle fermée, soit sensiblement une proportion 3/4 - 1/4. Ces proportions phénotypiques sont caractéristiques des résultats d'un croisement de deux parents de lignée pure différant par un caractère à deux allèles: l'un étant dominant et l'autre récessif. Pour ce second caractère, les deux allèles sont donc O = ouvert (dominant) et f = fermé (récessif).
b) Les génotypes des parents de race pure sont donc RR,OO X BB,ff ou réciproquement RR,ff X BB,OO puisque les deux caractères sont indépendants.
c) Quels que soient les génotypes des parents, les enfants F1 de parents homozygotes sont tous semblables et combinent les allèles des parents pour chaque caractère. Leur génotype RB,Of se traduit par le phénotype "fleur rose à corolle ouverte".
d) Si l'on construit l'échiquier permettant de déterminer tous les enfants F2 d'un couple de F1 RB,Of X RB,Of, on peut observer que les F2 à fleur rose et corolle ouverte peuvent être de deux génotypes différents: RB,OO ou RB, Of:
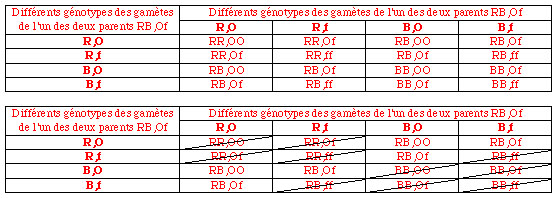
e) Les 165 plantes à fleur rose ouverte sont RB pour la couleur de la fleur, mais certaines sont OO, d'autres Of pour l'ouverture de la corolle. Afin de préciser, pour chacune d'elles, le génotype pour le second caractère, il convient d'effectuer, pour chacune d'elle, un testcross, c'est-à-dire de croiser chacune d'elles avec une plante homozygote récessive pour le second caractère, c'est-à-dire à corolle fermée.
Si la plante testée est Of, son croisement avec une plante ff donnera moitié de descendants à fleur ouverte (Of) et moitié à fleur fermée (ff), mais si la plante testée est OO, son croisement avec une plante ff donnera 100% de plantes à fleur ouverte (Of). Comme pour tout testcross, les proportions des différents phénotypes issus du croisement d'un individu de génotype inconnu avec un individu homozygote récessif pour un caractère déterminé, égalent les proportions des allèles correspondants composant le génotype de l'individu "testé".
D'après le tableau précédent, on s'attend à trouver 1/3 (2 cases sur les 6 de plantes à fleur rose) de plantes F2 de génotype RB,OO et 2/3 (4 cases sur les 6 de plantes à fleur rose) de plantes F2 de génotype RB,Of. Sur 165 plantes à fleur rose, on s'attend à trouver 55 plantes de génotype RB,OO et 110 plantes de génotype RB,Of.
***
Sachant qu'un couple constitué d'un homme A+ et d'une femme B- a un premier enfant A+, un deuxième A- et enfin un troisième B+, quelle était la probabilité que ces trois enfants apparaissent dans l'ordre indiqué?
Le couple ayant à la fois des enfants A et B, les parents A et B ne peuvent être que de génotype Ao et Bo. S'ils ont un enfants Rh-, c'est-à-dire de génotype rr, le père Rh+ ne peut être que Rr. Un tel couple Ao,Rr X Bo,rr a 1/8 de chance d'avoir un enfant Ao,Rr, 1/8 de chance d'avoir un enfant Ao,rr et 1/8 de chance d'avoir un enfant Bo,Rr.

La probabilité que ces trois enfants apparaissent dans l'ordre indiqué égale:
1/8 . 1/8 . 1/8 = 0,00195.
***
La maladie de Tay-Sachs ou idiotie familiaile amaurotique infantile (l’amaurose est une cécité sans altération de l'oeil ou du système optique) caractérise les enfants de génotype ii; ils meurent très jeunes. Un individu sain peut être aussi bien Ii que II. Par ailleurs, un individu brachydactyle (qui a les doigts courts) est de génotype NB, NN étant normal et BB létal. Quels sont les proportions des différents phénotypes parmi les enfants viables d’un couple Ii,NB X Ii,NB?
Il convient dans un premier temps de produire un échiquier donnant tous les génotypes -et leurs proportions, ici en seizièmes- attendus parmi les enfants de ce couple.
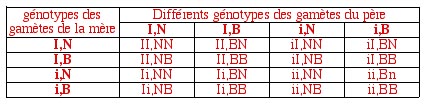
Ensuite, il convient de supprimer les enfants qui se sont pas viables, car présentant le génotype BB; les proportions des enfants viables s'epriment dès lors en douzièmes.
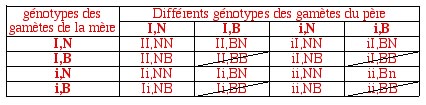
Il suffit enfin de traduire les génotypes en proportions phénotypiques.
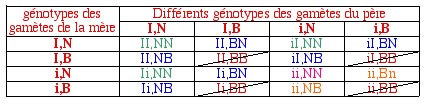
On obtient:
***
Génétique et sexualité
On croise un coq "Sussex" blanc avec une poule "Rhode Island" rouge. Tous les poussins issus de ce croisement sont blancs. On croise par ailleurs un coq "Rhode Island" rouge avec une poule "Sussex" blanche. On obtient dans ce cas 50% de poussins blancs et 50% de poussins rouges. Donnez le génotype d'un coq "Sussex" blanc et d'une poule "Rhode Island" rouge.
Puisque la fréquence des phénotypes des enfants d'un couple dépend du sexe des parents, il s'ensuit que le caractère envisagé est porté par une portion de chromosome sexuel présent chez un sexe seulement. Chez les poules, oiseaux gallinacés, les chromosomes sexuels sont ZZ chez le mâles et Z- chez la femelle.
On comprend qu'un coq blanc ZBZB donne avec une poule rouge Zr- 100% de poussins blancs, soit mâles ZBZr, soit femelles ZB-.
Par contre, le croisement réciproque d'un coq rouge ZrZr donne avec une poule blanche ZB- 50% de poussins mâles blancs ZBZr et 50% de poussins femelles rouges Zr-.
***
Gustave, hémophile -mais non daltonien- et dont le frère l’est également, convole en justes noces avec l’élégante Eglantine, qui ne présente aucun défaut. Ils ont 7 enfants : Pierre et Jacques, garçons daltoniens -mais non hémophiles-, Jean et Paul, qui n’ont aucun défaut, et trois filles sans problème : Gertrude, Cunégonde et Albertine. Vingt ans plus tard, Gertrude met au monde un petit Alfred normal, Albertine accouche d’un petit Jules hémophile -mais non daltonien- comme son grand-père et le bébé de Cunégonde est un mignon garçon, malheureusement daltonien -mais non hémophile-. Donnez le génotype d’Eglantine et de ses filles Gertrude et Cunégonde.
Il y a deux caractères portés par le chromosome X, l'un concernant le daltonisme, l'autre l'hémophilie, qu'il ne faut pas confondre, et dont on notera les allèles de la façon suivant: Xd = daltonien, XD = non daltonien, Xh= hémophile et XH = non hémophile.
Eglantine a donné deux types de chromosomes X à ses garçons: XdH à ses garçons Pierre et Jacques daltoniens et non hémophiles de génotype XdHY, et XDH à ses garçons Jean et Paul qui ne sont ni daltoniens ni hémophiles, donc de génotype XDHY. Eglantine présente donc un génotype XdHXDH. Si Gustave est non daltonien mais hémophile, son génotype est XDhY et il a donné son unique chromosome XDh à chacune de ses trois filles dont Gertrude et Cunégonde. Gertrude, ayant donné un chromosome XDH à son enfant mâle Alfred normal de génotype XDHY, est donc de génotype XDhXDH. Cunégonde ayant, quant à elle, donné un XdH à son garçon daltonien de génotype XdHY, présente le génotype XDhXdH.
***
Un gène récessif h lié au sexe retarde le temps de coagulation du sang et est responsable de l'hémophilie. André et Bernadette, non hémophiles, ont ensemble quatre enfants: Albert est sain, Bénédicte est saine, de même que Charlotte, mais Denis est hémophile.
a) Si Bénédicte se marie avec un homme normal, quelle est la probabilité que son premier enfant soit un garçon hémophile?: 1/8
Si André est non hémophile, son phénotype est nécessairement XHY puisque, comme tout homme, il ne possède qu'un allèle H pour ce caractère, porté par la portion du chromosome X non homologue à Y, et qui détermine son phénotype. Bernadette a nécessairement un allèle normal H, puisqu'elle est de phénotype non hémophile, mais aussi un allèle h puisqu'elle l'a donné Denis, son garçon hémophile. Bénédicte a nécessairement hérité de André un XH, mais elle a une (mal)chance sur deux d'avoir reçu une XH de sa mère Bernadette, et une chance sur deux d'en avoir hérité un Xh. Bénédicte est donc XHXH ou XHXh, de façon équiprobable. En faisant un garçon qui reçoit nécessairement Y de son mari, elle a une (mal)chance sur deux de lui donner XH et une (mal)chance sur deux de lui donner son second chromosome X, qui est soit XH soit Xh de façon équiprobable. Elle n'a donc qu'une (mal)chance sur quatre de donner un Xh à son enfant. En outre, elle a une chance sur deux de donner naissance à un garçon. La probabilité que son premier enfant soit un garçon hémophile vaut donc 1/4.1/2=1/8
b) Supposez que son premier enfant soit hémophile. Quelle est la probabilité que son second enfant soit un garçon hémophile?: 1/4
Si le première enfant de Bénédicte est hémophile et comme son mari ne l'est pas, on est certain qu'elle porte un Xh, et qu'elle est donc de génotype XHXh. Elle a donc qu'une (mal)chance sur deux de donner un Xh à son enfant. En outre, elle a une chance sur deux de donner naissance à un garçon. La probabilité que son premier enfant soit un garçon hémophile vaut donc 1/2.1/2=1/4
c) Si Charlotte se marie avec un homme hémophile, quelle est la probabilité que son premier enfant soit normal?: 3/4
Charlotte a nécessairement hérité de André un XH, mais elle a une (mal)chance sur deux d'avoir reçu une XH de sa mère Bernadette, et une chance sur deux d'en avoir hérité un Xh. Bénédicte est donc XHXH ou XHXh, de façon équiprobable. Elle a donc 3 chances sur 4 de transmettre à son enfant l'allèle H. Ce que son mari transmet à ses enfants n'a aucune importance, puisqu'il s'agit soit d'un chromosome Y qui ne porte aucun allèle pour ce caractère, soit un Xh puisqu'il est daltonien et que, dans ce cas, l'allèle h ne permet pas de produire un enfant non hémophile.
d) Si la mère de Bernadette était phénotypiquement normale, quel était le phénotype de son père? On ne peut rien affirmer à son sujet, car Bernadette est de génotype XHXh et l'on ne sait si c'est XH ou Xh qu'elle a reçu de sa mère normale mais peut-être porteuse. Dès lors, on ne sait si c'est XH ou Xh qu'elle a reçu de son mère père, qui aurait pu être normal ou hémophile.
***
Caractère influencé par le sexe mais non porté par un gonosome
La calvitie est un caractère influencé par le sexe (mais non porté par le chromosome X ni par le chromosome Y) qui est dominant chez l'homme et récessif chez la femme. Sur un échantillon de 10000 hommes, 7225 n'étaient pas chauves. Combien de femmes chauves peut-on s'attendre à trouver sur un échantillon de 10000 femmes ?
Le caractère présente l'allèle n normal dominant chez la femme mais récessif chez l'homme, et l'allèle c chauve dominant chez l'homme et récessif chez la femme.
On sait que le génotype des hommes normaux est nn. De tels hommes apparaissent donc au sein d'une population lorsque chacun des parents donne à l'enfant l'allèle n. La probabilité d'apparition d'un homme normal vaut donc le carré de la fréquence d'apparition de l'allèle n (un peu comme la fréquence d'apparition d'un 22 en jetant deux dés à six faces équiprobables vaut 1/6.1/6 = 1/36). On calcule dès lors la fréquence de l'allèle n dans la population en prenant la racine carrée de la fréquence des hommes normaux de 7225/10000 = 0,7225.
La fréquence de l'allèle normal, récessif chez l'homme mais dominant chez la femme, vaut donc: (0,7225)1/2 = 0,85
La fréquence de l'allèle chauve c, dominant chez l'homme mais récessif chez la femme, vaut donc: 1 - ,085 = 0,15
Le génotype des femmes chauves est cc.
La fréquence d'apparition des femmes chauves vaut donc: 0,152 = 0,0225
Le nombre attendu de femmes chauves sur 10 000 femmes est de: 0,0225 . 10000 = 225
***
Epistasie
On croise deux lignées pures de pois blancs et l'on obtient en F1 100% de pois colorés pourpres. Combien de pois blancs et de pois colorés obtiendra-t-on théoriquement sur 100 F2?
L'expression de la couleur des pois peut être inhibée à différents niveaux de sa synthèse, et la couleur blanche peut être obtenue par différentes mutations. Définissons théoriquement deux caractères A et B dont l'expression normale du gène est nécessaire pour la synthèse du colorant pourpre: un pois n'est pourpre que s'il possède au moins un allèle normal dominant pour chacun de ces deux caractères. Si les lignées parentales sont pures, elle sont homozygotes, et, sachant que leur croisement donne des pois colorés, elles ne sont pas mutées au niveau du même caractère. Par contre, la mutation de chacune d'elle est compensée par un allèle normal provenant de l'autre parent: c'est un cas d'épistasie. Le croisement parental AA,bb X aa,BB donne 100% de Aa,Bb, tous colorés. Le croisement des F1 Aa,Bb X Aa,Bb donne 9/16 de F2 possédant au moins un "A" et un "B", donc colorés, et 7/16 de blancs. Sur 100 individus cela donne 56 plantes à pois colorés et 44 blancs.
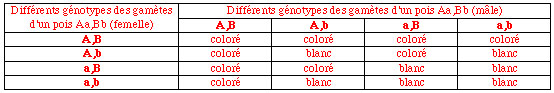
***
Génétique et probabilité
Quelle est la probabilité, pour un couple constitué d'un homme daltonien et d'une femme phénotypiquement normale mais porteuse d'un allèle daltonien, d'avoir un garçon daltonien, puis une fille normale, puis enfin un garçon normal?
On commence par déterminer la probabilité d'apparition de chaque phénotype demandé, pris séparément, parmi les enfants possibles d'un tel couple, en considérant tous les génotypes possibles.

On en déduit que ce couple aura un garçon daltonien avec 25% de chance, un garçon normal avec 25% de chance, une fille daltonienne avec 25% de chance et une fille normale avec 25% de chance.
Ensuite, on détermine la probabilité, pour ce couple, d'avoir successivement et dans l'ordre, un garçon daltonien (0,25), puis une fille normale (0,25), puis enfin un garçon normal (0,25):
0,25.0,25.0,25 = 0,253 = 0,02 = 2%
***
Sachant que l'allèle récessif d, déterminant le daltonisme, est porté par une portion du gonosome (= chromosome sexuel) X non homologue à Y, et qu'il y a 1 femme sur 900 qui est daltonienne, calculez la probabilité, pour un couple constitué par un homme daltonien et par une femme phénotypiquement normale, d'avoir, sur deux enfants, une fille normale et un garçon normal.
Il convient d'abord, de calculer la fréquence d'apparition des allèles d et D, tous deux portés par le chromosome X. On sait qu'une femme daltonienne est nécessairement XdXd. Une telle femme apparaît donc au sein d'une population lorsque chacun des parents donne à l'enfant l'allèle Xd. La probabilité d'apparition d'une femme daltonienne vaut donc le carré de la fréquence d'apparition de l'allèle Xd (un peu comme la fréquence d'apparition d'un 33 en jetant deux dés à six faces équiprobables vaut 1/6.1/6 = 1/36). On calcule dès lors la fréquence de l'allèle Xd dans la population en prenant la racine carrée de la fréquence des femmes daltoniennes:
(1/900)1/2 = 1/30 = 0,033
Comme il n'existe que deux allèles pour ce caractère, l'allèle normal XD (non daltonien) apparaît donc avec une fréquence de
1 - 0,033 = 0,967
On observe ainsi, parmi toutes les femmes de la population, trois génotypes possibles:
On vérifie, par souci de vérifier que l'on a oublié aucun génotype, que la somme des fréquences de tous les phénotypes possibles vaut bien 100%: 0,935 + 0,064 + 0,001 = 1,000
On sait (certitude de l'énoncé) que la femme du couple est normale et ne peut donc présenter comme génotype que XDXD ou XDXd.
L'homme du couple ne peut être daltonien (énoncé) qu'en présentant le génotype XdY.
Déterminons, pour chacun des couples possibles, la fréquence d'apparition des différents phénotypes parmi les enfants F1:


Pour répondre à la question et pour chacun des deux couples possibles, deux cas de figure sont possibles: soit la fille normale naît avant le garçon normal, soit l'inverse: on multiplie donc le produit des probabilités d'avoir un garçon norma et une fille normale par deux. Le produit de ces probabilité est à pondérer, pour chaque couple, par la probabilité d'apparition de ce couple. On additionne enfin les probabilités des différents cas de figure qui répondent à la demande de l'énoncé du problème:
0,936 . (2 . 2/4 . 2/4) + 0,064 . (2 . 1/4 . 1/4) = 0,476 ou 47,6%
***
Un radis peut être long (LL), rond (RR) ou ovale (LR). Des plantes à radis longs sont croisées avec des plantes à radis ovales. Les enfants F1 ainsi produits sont croisés au hasard. Donnez les proportions des différents phénotypes obtenus en F2.
Lors du croisement LL X LR, LL donne 100% de gamètes L et LR donne 50% de gamètes L et 50% de gamètes R. La génération F1 est donc constituée de 50% de plantes à radis longs LL et de 50% de plantes à radis ovales LR.
Si les individus obtenus en F2 sont le résultat de croisements aléatoires entre individus F1, ils sont le produit de 4 croisements possibles, dont les probabilités d'apparition sont les suivantes:
On détermine, en construisant si nécessaire un échiquier de croisement, pour chacun des couples, la fréquence théorique d'apparition de chaque génotype (et donc phénotype) en F2:
On additionne enfin ces probabilités d'apparition des différents phénotypes F2, bien sûr pondérées par les probabilités respectives d'apparition des couples qui leur ont donné naissance. On obtient donc en F2:
La somme de ces fréquences donnant bien 100%, on n'a oublié aucun cas.
***
Chez le lapin, la maladie de Pelger se traduit par une anomalie de la division des leucocytes. Les individus atteints sont Pp, les individus normaux sont PP et les individus pp meurent à la naissance. Si des individus atteints sont croisés entre eux et que l'on croise au hasard les F1 produits, quels seront, parmi les lapins viables, les différents phénotypes et leur fréquence d'apparition en F2?
La génération F1 est le résultat du croisement Pp X Pp et est donc constituée de 25% de PP, 50% de Pp et 25% de pp:

Mais parmi les lapins F1, les pp meurent à la naissance et il reste, à l'âge de la reproduction, 1/3 de PP et 2/3 de Pp:

Si les individus obtenus en F2 sont le résultat de croisements aléatoires entre individus F1, ils sont le produit de 4 croisements possibles, dont les probabilités d'apparition sont les suivantes:
On détermine, en construisant si nécessaire un échiquier de croisement, pour chacun des couples, la fréquence théorique d'apparition de chaque génotype (et donc phénotype) en F2:
On additionne enfin ces probabilités d'apparition des différents phénotypes F2, bien sûr pondérées par les probabilités respectives d'apparition des couples qui leur ont donné naissance. On obtient donc en F2:
***
Connaissant les risques d'immunisation fœto-maternelle vis-à-vis de l'agglutinogène Rh de la part d'une femme rhésus- portant un enfant rhésus+, et sachant que 85% des gens de notre population sont rhésus+,
a) calculez la probabilité d'apparition d'un couple homme rhésus+ X femme rhésus-;
b) calculez, pour ce couple, la probabilité d'avoir un enfant rhésus+ suivi d'un autre rhésus+ et donc de connaître un risque d'hémolyse du sang du second enfant.
a) Si 85% des gens rhésus+ sont RR ou Rr, on en déduit que 15% des gens sont rhésus- rr. On sait qu'une personne rhésus- est nécessairement rr. Une telle personne apparaît donc au sein d'une population lorsque chacun des parents donne à l'enfant l'allèle r. La probabilité d'apparition d'une personne rhésus- vaut donc le carré de la fréquence d'apparition de l'allèle r (un peu comme la fréquence d'apparition d'un 44 en jetant deux dés à six faces équiprobables vaut 1/6.1/6 = 1/36). On calcule dès lors la fréquence de l'allèle r dans la population en prenant la racine carrée de la fréquence des personnes rhésus-:
(0,15)1/2 = 0,39
L'allèle R présente donc une fréquence de 1 - 0,39 = 0,61
Les différents génotypes apparaissent donc avec des fréquences de:
Deux types de couples répondent à la question de l'énoncé et peuvent connaître un risque d'hémolyse du sang du second enfant: homme RR X femme rr et homme Rr X femme rr. Il apparaissent avec une fréqunce de:
Un couple à risque a donc 5,6% + 7,2% = 12,8% de chance de voir le jour dans notre population.
b) On sait (certitude de l'énoncé) que l'homme du couple est rhésus+ et ne peut donc présenter comme génotype que RR ou Rr.
La femme ne pouvant être rhésus- que par son génotype rr, il apparaît deux types de couples répondant à l'énoncé:
On détermine, pour chacun de ces couples, la probabilité de concevoir un enfant rhésus+:


Pour le premier couple, tous les enfants sont rhésus+, pour le second seulement 50% des enfants sont rhésus+.
La probabilité pour un couple homme rhésus+ X femme rhésus- d'avoir deux enfants rhésus+ est donc la somme de ces probabilités, pour chacun des couples possibles, pondérées par la fréquence d'apparition théorique des couples respectifs:
100% . 100% . 44% + 50% . 50% . 56% = 58%
***
Sachant que le phénotype albinos, gouverné par un allèle récessif, apparaît, dans une population de lapins, avec une fréquence de 1 animal sur 1000, calculez la probabilité, pour un couple de lapins choisis au hasard, d'avoir un premier petit albinos suivi d'un second normal.
Il existe trois génotypes possibles: AA (lapin normal), Aa (lapin normal) et aa (lapin albinos). Si les lapins aa apparaissent avec une fréquence de 1/1000 au sein d'une population lorsque chacun des parents donne à l'enfant l'allèle a, la probabilité d'apparition d'un lapin albinos vaut donc le carré de la fréquence d'apparition de l'allèle a (un peu comme la fréquence d'apparition d'un 55 en jetant deux dés à six faces équiprobables vaut 1/6.1/6 = 1/36). On calcule dès lors la fréquence de l'allèle a dans la population en prenant la racine carrée de la fréquence des lapins albinos:
(1/1000)1/2 = 0,032
L'allèle A présente donc une fréquence de 1 - 0,032 = 0,968
Les différents génotypes apparaissent donc avec une fréquence de:
Parmi les différents couples de lapins possible formés au hasard, seuls les suivants peuvent avoir un petit albinos et un autre normal:
En multipliant, pour chacun des couples, la probabilité d'avoir un premier petit albinos suivi d'un second petit normal, par la fréquence théorique d'apparition du couple correspondant, la réponse est:
50% . 50% . 1,22 . 10-4 + 75% . 25% . 0,004 = 0,0008 soit 0,08%, ce qui n'est pas grand'chose.
***