
Les lois de Mendel
1 Introduction et terminologie
Au cours des générations successives, les parents transmettent des caractères à leurs descendants (ceux qui permettent entre autres de reconnaître l'espèce à laquelle l'enfant appartient), ces caractères héréditaires pouvant être morphologiques (comme la taille, la couleur de la peau, des cheveux ou des yeux, la forme de certains organes comme le nez ou le lobe des oreilles), physiologiques (comme la résistance aux maladies, la sensibilité olfactive à l'androstérone présente dans la sueur et l'urine des mammifères mâles), biochimiques (comme la phénylcétonurie, une affection héréditaire empêchant la dégradation de l'acide aminé "phénylalanine", la capacité de dégrader l'alcool ingéré) ou éthologiques -c'est-à-dire comportementaux- (de nombreux comportements innés ont été étudiés chez les animaux, comme l'imprégnation ou l'émission du chant chez certaines espèces d'oiseaux).
Rappelons à ce propos que les maladies héréditaires sont dues aux gamètes portant un patrimoine héréditaire altéré, et non à une altération de l'oeuf, de l'embryon ou du foetus, ce qui provoque des maladies congénitales non héréditaires. Il n'est pas rare de voir dans une même famille réapparaître un caractère qui ne s'était plus manifesté pendant une ou plusieurs générations: on parle alors d'atavisme et de caractère atavique.
L'hérédité est donc l'ensemble des caractères passant des ascendants aux descendants. La génétique est la science qui étudie l'hérédité et la transmission des caractères héréditaires. Les caractères de l'enfant, ressemblant à ceux du père, à ceux de la mère ou intermédiaires, sont déterminés par le phénomène de fécondation : les faits d'observation et d'expérimentation ne sont compréhensibles que si les caractères héréditaires sont supportés à la fois par l'ovule et le spermatozoïde.
Cette science génétique a connu trois périodes d'évolution importantes :
2 Expériences d'hybridation
2.1 Premières tentatives
L'hybridation est la principale méthode expérimentale permettant d'étudier la transmission des caractères héréditaires. La méthode d'hybridation fut appliquée pour la première fois en 1826 par Sageret et de 1859 à 1863 par le botaniste français Naudin. Malheureusement, les espèces sur lesquelles travaillait Naudin différaient par beaucoup de caractères. L'analyse des hybrides était si difficile que Naudin ne parvint pas à énoncer les lois de la transmission héréditaire.
2.2 Les lois de Mendel
2.2.1 Circonstances historiques
Johann Mendel (1822-1884), dit Gregor en religion, moine botaniste autrichien, mit le premier clairement en évidence les lois de la transmission de l'hérédité. C'est dans les jardins du monastère des Augustins à Brno, en Moravie, qu'il s'adonna à la culture et l'hybridation des pois. Il publia en 1866 le résultat de ses expériences, mais ce travail fondamental passa inaperçu. Il fallut attendre 1900 pour que son ouvrage soit révélé au public des biologistes. A cette date, les lois de l'hybridation, telles que les avait formulées Mendel, furent redécouvertes indépendamment par trois botanistes : de Vries aux Pays-Bas, Correns en Allemagne et von Tschermak en Autriche. A partir de 1902, Cuénot en France et Bateson en Angleterre généralisaient ces lois aux animaux.
2.2.2 Monohybridisme
Une lignée pure est un ensemble de générations successives semblables pour tous les caractères ; les enfants sont donc identiques en tous points aux parents.
Le croisement de deux individus appartenant à deux lignées pures ne différant que par un seul caractère constitue un cas de monohybridisme.
Si l'on croise à la génération parentale "P" une fleur rouge et une fleur blanche de Mirabilis jalapa, la Belle de nuit, on obtient en première génération "F1" des hybrides roses tous semblables.
La première loi de Mendel, ou loi d'association, dit que "les hybrides issus du croisement de parents de lignée pure différant par un caractère sont tous semblables et associent les caractères parentaux", ici les couleurs rouge et blanches.

Ketmies de Syrie Hibiscus syriacus, Malvaceae,Malvales, Magnoliopsides (=Dicotylédones): à gauche, la variété 'Blue Bird', à droite la variété 'Woddbridge' et au centre l'hybride obtenu, de couleur intermédiaire entre les parents et qui associe donc les caractères parentaux. (Jardin à Hamois, Belgique - 11/08/2005 - Photographies originales réalisées par Eric Walravens).
En réalité, le caractère "couleur de la fleur" est représenté chez les parents par deux allèles, respectivement "rouge" et "blanc". Et il aisé de comprendre pourquoi tous les F1, recevant l'allèle "rouge" d'un parent et l'allèle "blanc" de l'autre, les associent pour paraître à la fois rouges et blancs, c'est-à-dire roses.
Si l'on croise deux de ces fleurs roses F1, à la seconde génération "F2", on obtiendra des plantes à fleurs rouges, d'autres à fleurs blanches et d'autres à fleurs roses. Si l'expérience a été faite sur un grand nombre d'individus, on constate que ces différents types apparaissent selon des proportions définies : 1/4 de plantes à fleurs rouges, 1/4 de plantes à fleurs blanches et 2/4 de plantes à fleurs roses.
La deuxième loi de Mendel, ou loi de disjonction, dit qu' "au départ d'un croisement entre individus de lignée pure différant par un caractère, les allèles associés à la première génération se disjoignent à la seconde génération, avec retour de la moitié des individus au type des grands-parents".
Afin de comprendre ces faits, il faut se rappeler que les plantes et les animaux présentent, à l'état adulte reproducteur, dans chacune de leurs cellules diploïdes, deux jeux de chromosomes, autrement dit deux fois l'ensemble des caractères héréditaires: l'un vient du père, l'autre de la mère, chacun des parents ne donnant qu'un gamète haploïde. Associés dès la fécondation, ce sont ces deux patrimoines qui déterminent l'hérédité de l'individu. Ainsi, pour chaque caractère, tout organisme possède deux allèles, semblables ou différents: c'est le génotype. On a l'habitude de représenter, pour chaque caractère, tout allèle par une lettre, et donc chaque génotype par deux lettres: "RR" signifie, dans notre exemple, que les cellules d'une plante à fleurs rouges portent l'allèle "fleurs rouges" sur chacun de leurs deux jeux de chromosomes, "RB" nous apprenant pour une plante à fleurs roses que chaque cellule possède un allèle "fleurs blanches" et un autre "fleurs rouges", chacun provenant d'un des parents de la plante à fleurs roses. On parle, pour un caractère déterminé, de génotype homozygote si les deux allèles sont identiques et de génotype hétérozygote s'ils sont différents.
Le phénotype est l'expression extérieurement visible du génotype, c'est-à-dire de l'association des deux allèles pour le caractère considéré.
Nos deux croisements successifs peuvent se résumer ainsi:

2.2.3 Dominance et récessivité
Dans l'exemple précédent, les allèles B et R ont une importance équivalente dans la détermination du phénotype floral: on dit qu'ils sont codominants. Mais ce n'est pas toujours le cas. Formons par exemple, à partir de deux lignées pures de souris, l'une à pelage blanc, l'autre à pelage gris foncé, des couples d'un mâle gris et d'une femelle blanche ou inversement (les résultats n'en seront pas changés). Tous les enfants F1 auront le pelage gris foncé, et en F2, les 3/4 seront gris et 1/4 sera blanc! Il est facile d'expliquer ces résultats si l'on considère que l'allèle blanc "b" ne s'exprime pas en présence de l'allèle "G" qui lui est dominant.
On dira qu'un allèle est récessif s'il ne s'exprime pas en présence d'un allèle dominant. C'est le cas pour des allèles correspondant à un gène ne traduisant aucune protéine, donc à l'absence du caractère en question: l'albinisme (absence de mélanine colorant la peau), le daltonisme (incapacité de déceler certaines couleurs et confusion entre couleurs), l'hémophilie (maladie hémorragique liée au déficit de facteurs de coagulation dans le plasma sanguin) sont quelques exemples d'allèles récessifs pour des caractères pigmentaire, visuel ou sanguin.
On prendra l'habitude de représenter un allèle récessif par une lettre minuscule et un allèle dominant par une lettre majuscule. Dans notre exemple :


Polémoine Polemonium caeruleum, Polemoniaceae, Polemoniales, Spermatophytes. Chez cette espèce, le caractère "couleur de la fleur" est représenté par deux allèles: bleu "B" est dominant et "b" blanc est récessif. Le génotype de la plante illustrée est donc soit "BB", soit "Bb". (Jardin alpin du col du Lautaret, Ecrins, France - 20/07/1990 - Diapositive originale réalisée par Eric Walravens).
2.2.4 Dihybridisme
Les expériences de dihybridisme consistent à croiser deux individus de lignée pure différant par deux caractères. Mendel a croisé des plantes donnant des pois jaunes et lisses avec des plantes à pois verts et ridés. En F1, il a obtenu 100 % de plantes donnant des pois jaunes et lisses, et en F2 il a compté 9/16 de pois jaunes et lisses, 3/16 de pois jaunes et ridés, 3/16 de pois verts et lisses et 1/16 de pois verts et ridés. On comprend aisément ces résultats si l'on considère que "jaune" est un allèle dominant et "vert "un allèle récessif pour le premier caractère "couleur du pois", et d'autre part que "lisse" est dominant et "ridé" récessif pour le second caractère.
En partant des génotypes homozygotes connus des parents JJ,LL et vv,rr et sachant que tout gamète est haploïde (c'est-à-dire qu'il ne possède qu'un allèle par caractère) à l'issue de la réduction chromatique de la méiose ayant lieu au cours de la gamétogenèse, on construit un "échiquier de Punnet" où l'on inscrit l'haplotype (= génotype haploïde) des différents gamètes que chaque géniteur donne et où l'on en déduit la proportion des différents génotypes -et donc des fréquences des différents phénotypes- des enfants. Dans notre exemple, cela donne pour la première génération:

...et pour la seconde génération F2, issue de parents F1 :

Cette fois, les résultats sont très différents du monohybridisme: dans la descendance F2 des hybrides F1 croisés entre eux, on constate non seulement la réapparition d'associations primitives d'allèles (pois jaunes et lisses, pois verts et ridés), mais aussi la présence de phénotypes constituant de nouvelles associations (pois jaunes et ridés, pois verts et lisses). Les génotypes correspondants sont constitués entre autres par l'association de deux couples d'allèles qui appartenaient l'un et l'autre aux génotypes initiaux de la génération P: JJ,rr et vv,LL. C'est l'illustration de la troisième loi de Mendel ou loi de ségrégation des allèles qui dit que "lorsqu'on croise deux lignées pures différant par plusieurs caractères, on voit apparaître, à partir de la seconde génération, des lignées différentes présentant des nouvelles combinaisons de caractères par rapport à celles existant chez les lignées de départ".
Un autre cas de dihybridisme est celui de la robe du bœuf domestique Bos taurus, dont l'aspect peut être uni ou tacheté (="pie") et la couleur noire ou rousse: si l'on croise un taureau pie noir de lignée pure avec une vache à la robe unie et rousse de lignée pure, les veaux produits sont tous unis et noirs. On en déduit que l'allèle "robe unie" est dominant par rapport à l'allèle "pie", et que, d'autre part, l'allèle "noir" domine l'allèle "roux" récessif. Si ces enfants sont croisés entre eux, il apparaît en seconde génération 9/16 de bœufs unis et noirs, 3/16 de bœufs pies noirs, 3/16 de bœufs unis et roux et 1/16 de boeufs pies roux, cette dernière combinaison phénotypique apparaissant pour la première fois.
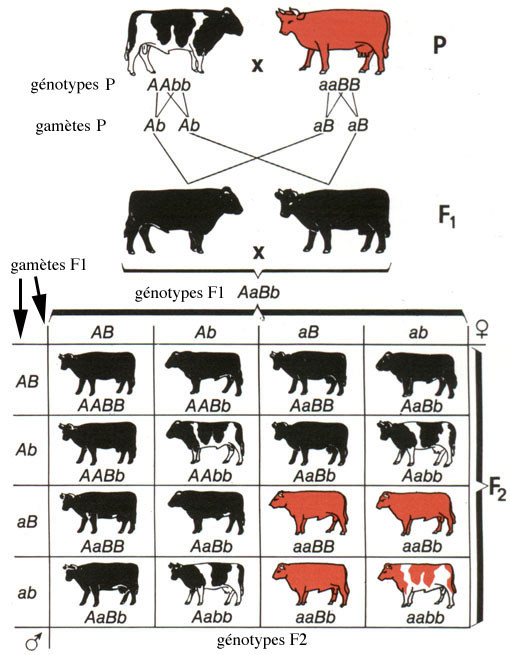
Tableau représentant deux générations de bœufs issus du croisement d'un taureau pie noir de lignée pure avec une vache à la robe unie et rousse de lignée pure (d'après Fast, 1972, traduit).
Les trois lois de Mendel sont vérifiées pour le polyhybridisme, relatif à des croisements entre lignées différant par plus de deux caractères.
2.3 Caractère gouverné par plus de deux allèles
Il existe beaucoup de caractères qui se présentent sous des phénotypes multiples, chacun étant gouverné par un allèle différent. Nous en verrons deux exemples.
2.3.1 La coloration du pelage des lapins
La coloration du pelage des lapins peut se présenter sous trois aspects :
En croisant des lignées pures de lapins sauvages et albinos, les hybrides F1 hétérozygotes présentent le phénotype sauvage : l'allèle "sauvage" domine donc l'allèle "albinos". Parallèlement, en croisant des lapins "albinos" et "Himalaya" de lignées pures, les F1 sont de type "Himalaya". Si l'on accouple enfin les F1 "sauvage" avec les F1 "Himalaya", on obtient 50 % d'hétérozygotes de type "sauvage", 25 % d'hétérozygotes de type "Himalaya" et 25 % de phénotypes "albinos", nécessairement homozygotes.
Cette expérience montre que l'allèle "albinos" est récessif par rapport à l'allèle "Himalaya", celui-ci se comportant comme récessif vis-à-vis de l'allèle dominant "sauvage".

2.3.2 Le système sanguin ABo
Les différents groupes sanguins correspondent aux différents antigènes que les hématies peuvent porter à leur surface. En ce qui concerne le système ABO, une personne est du groupe sanguin A si ses globules rouges portent l'antigène A (45% des personnes en Belgique), du groupe sanguin B si ses globules rouges portent l'antigène B (8% des personnes en Belgique), du groupe sanguin AB si ses globules rouges portent à la fois les antigènes A et B (3% des personnes en Belgique), et du groupe sanguin O ("O" pour "ohne"= "sans" en allemand) si ses globules rouges ne portent aucun antigène (44% des personnes en Belgique).
Comme tout être humain fabrique des anticorps dressés contre tout antigène qui lui est étranger, une personne du groupe A fabrique des anticorps anti-B dressés contre l'antigène B, une personne du groupe B fabrique des anticorps anti-A dressés contre l'antigène A, une personne du groupe O fabrique à la fois des anticorps anti-A et anti-B dressés respectivement contre les antigènes A et B, et une personne du groupe AB ne fabrique aucun anticorps dressé contre A ou B.
Lorsque les anticorps rencontrent les antigènes correspondants, il s'ensuit une réaction d'agglutination, agglutinant, c'est-à-dire collant en masse les globules rouges. Pour cette raison, les antigènes qui nous occupent sont appelés agglutinogènes et les anticorps agglutinines. C'est le pathologiste autrichien K. Landsteiner qui découvrit, en 1900 et 1901, les différents groupes sanguins du système ABO, en mélangeant les sangs de diverses personnes et en observant les éventuelles réactions d'agglutination.

Hémogramme ABo de 3 élèves de l'AFB, chez qui 3 gouttes de sang ont été stérilement prélevées et mélangées respectivement à un sérum anti-A (universellement artificiellement coloré en bleu), à un sérum anti-B (universellement artificiellement coloré en jaune) et à un sérum anti-A et anti-B (universellement non coloré). L'éventuelle agglutination des hématies, formant des masses foncées sur le fond clair coloré du sérum, révèle la présence de l'antigène correspondant et indique l'appartenance du sang au groupe en question. Merci à Ludovic P. et à sa Maman pour les sérums fournis.
Si l'on transfuse une personne du groupe A avec du sang de groupe B, cette erreur peut provoquer deux réactions d'agglutination: d'une part, les agglutinogènes du donneur (Ag B) seront agglutinés par les agglutinines du receveur (Ab anti-B), d'autre part les agglutinogènes du receveur (Ag A) seront agglutinés par les agglutinines du donneur (Ab anti-A).
Si ces deux réactions ont lieu effectivement dans le cas d'une transfusion massive, seule la première réaction aura de graves conséquences en cas de transfusion modérée. En effet, une petite quantité de sang transfusé comporte beaucoup d'hématies et peu d'anticorps: la faible quantité d'agglutinines du donneur, diluée dans le sang du receveur, ne risque pas d'agglutiner beaucoup d'hématies du receveur, mais tous les globules rouges reçus seront agglutinés par les anticorps du receveur, provoquant dans ses vaisseaux sanguins capillaires des caillots induisant de graves accidents circulatoires.
On peut ainsi expliquer les divers cas d'incompatibilité de transfusion :
|
Groupe sanguin |
Agglutinogène(s) |
Agglutinine(s) |
Transfusion(s) possible(s) de sang : |
Transfusion(s) possible(s) de ce sang vers : |
|
A |
Ag A |
Ab anti-B |
A et O |
A et AB |
|
B |
Ag B |
Ab anti-A |
B et O |
B et AB |
|
AB |
Ag A et Ag B |
aucun |
A, B, AB et O |
AB |
|
O |
aucun |
Ab anti-A et anti-B |
O |
A, B, AB et O |
On appelle les gens du groupe O des "donneurs universels" car ils pourraient donner du sang, en faible quantité, à des gens de n'importe quel groupe, et on appelle les gens du groupe AB des "receveurs universels" car ils pourraient recevoir du sang des gens de n'importe quel groupe. Hormis les cas d'extrême urgente, on respectera néanmoins les groupes sanguins pour toute transfusion (sang A pour un transfusé A, B pour B,...). On a donc remplacé les anciennes notions de "donneur universel" et de "receveur universel" par celle de "transfusions isogroupes".
Les agglutinines anti-A et anti-B sont dites naturelles, dans la mesure où elles existent sans immunisation préalable (c'est-à-dire sans être générées par un premier contact avec l'antigène).
Génétiquement, l'allèle A code pour la synthèse de l'agglutinogène A et l'allèle B code pour la synthèse de l'agglutinogène B. Ce sont donc les deux allèles codominants. A côté d'eux, l'allèle récessifs o correspond à l'absence d'agglutinogène pour ce groupe sanguin. Sachant que toute personne est diploïde, les quatre phénotypes possibles peuvent correspondre aux génotypes suivants:
|
Phénotype A |
Phénotype B |
Phénotype AB |
Phénotype O |
|
AA |
BB |
AB |
oo |
|
Ao |
Bo |
|
|
2.3.3 Le système rhésus
Bien que le caractère Rhésus ne soit gouverné que par deux allèles, nous l'étudions ici afin d'associer son étude à celle du caractère ABo.
Les hématies du singe asiatique Macacus rhesus portent un agglutinogène baptisé "rhésus". Et 85 % des êtres humains portent sur leurs globules rouges cet Ag Rh : ils appartiennent au groupe sanguin Rh+, les 15 % restants ne portant pas cet antigène et étant baptisés Rh-.
Contrairement aux agglutinines anti-A et anti-B naturelles, l'agglutinine anti-Rh n'apparaît dans un sang Rh- qu'après un premier contact avec l'antigène Rh, c'est-à-dire après immunisation par l'antigène Rh. Cette immunisation apparaît:
Génétiquement, il existe deux allèles: l'un, dominant et baptisé R, code pour l'agglutinogène rhésus, l'autre, récessif et baptisé r, correspond à l'absence d'agglutinogène pour ce groupe sanguin.
2.3.4 Combinaison des systèmes ABo et rhésus
Parmi les 26 groupes sanguins connus, ABo et rhésus sont particulièrement importants quant aux transfusions possibles ou dangereuses. Ces deux caractères étant indépendants, on détermine le tableau suivant liant phénotypes et génotypes:
|
Phén. A+ |
Phén. A- |
Phén. B+ |
Phén. B- |
Phén. AB+ |
Phén. AB- |
Phén. o+ |
Phén. o- |
|
AA,RR |
AA,rr |
BB,RR |
BB,rr |
AB,RR |
AB,rr |
oo,RR |
oo,rr |
|
AA,Rr |
Ao,rr |
BB,Rr |
Bo,rr |
AB,Rr |
|
oo,Rr |
|
|
Ao,RR |
|
Bo,RR |
|
|
|
|
|
|
Ao,Rr |
|
Bo,Rr |
|
|
|
|
|

|
A+ |
A- |
B+ |
B- |
AB+ |
AB- |
o+ |
o- |
Total |
|
|
Personnes |
202 |
37 |
75 |
27 |
33 |
4 |
221 |
54 |
653 |
|
Pourcentage |
31% |
6% |
11% |
4% |
5% |
1% |
34% |
8% |
100% |
Histogramme original présentant la fréquence des groupes sanguins dans la population bruxelloise. L'échantillon est constitué de 653 personnes, élèves et parents d'élèves de l'Athénée Fernand Blum de Schaerbeek. 81% des personnes sont de groupe "Rhésus +" et 19% sont du groupe "Rhésus -".