
La méiose et la gamétogenèse
1 Nécessité de la méiose
Du point de vue chromosomique, la fécondation est un phénomène très simple: elle aboutit, à la suite de la fusion nucléaire de l'ovule et du spermatozoïde chez les animaux, de l'o(v)osphère et de l'anthérozoïde chez les plantes, à l'addition, dans le noyau du zygote, des jeux de chromosomes de chacun des gamètes. En bref, la fécondation d'une cellule haploïde par une autre cellule haploïde donne une cellule diploïde: n+n=2n (n est le nombre de chromosomes d'un simple jeu chromosomique, contenu dans une cellule haploïde). Chez l'Homme par exemple, les gamètes haploïdes (=cellules germinales) comportent chacun un jeu de 23 chromosomes (n=23) alors que le zygote diploïde issu de la fécondation (ainsi que toutes les cellules somatiques provenant des mitoses successives du zygote) comporte deux jeux de 23 chromosomes, soit 46 chromosomes en tout (2n=46).
Au contraire, le processus de méiose est un phénomène plus délicat puisqu'il doit permettre, à partir d'une cellule diploïde, d'obtenir des cellules haploïdes (des gamètes ou des spores par exemple), afin que, à la fécondation suivante, le nombre de chromosomes de l'espèce reste constant. Non seulement chaque cellule haploïde doit recevoir n chromosomes (des 2n chromosomes de la cellule qui lui a donné naissance), mais elle doit recevoir précisément un chromosome de chaque paire de chromosomes homologues. Contrairement à la mitose, mécanisme par lequel une cellule, haploïde ou diploïde, donne naissance à deux cellules-filles identiques, respectivement haploïdes ou diploïdes, la méiose est une série de deux divisions cellulaires successives par lesquelles une cellule diploïde donne naissance à quatre cellules haploïdes. Ces deux divisions successives sont désignées "méiose 1" ou division réductionnelle et "méiose 2" ou division équationnelle, et il ne s'effectue qu'une seule duplication de chromosomes avant la méiose 1.
2 Description de la méiose
2.1 La méiose 1 ou division réductionnelle
La méiose 1 est précédée par une interphase au cours de laquelle chacun des chromosomes se réplique et se retrouve constitué de deux chromatides-sœurs, parfaites copies l'une de l'autre.
Il s'ensuit une longue prophase 1, au cours de laquelle apparaissent les chromosomes homologues doubles de chaque paire apparaissent sous forme de longs filaments (stade leptotène), s'alignent (stade zygotène) puis se raccourcissent, s'épaississent (stade pachytène) et s'associent en une structure faite de quatre chromatides-sœurs appelée tétrade: les chromosomes homologues semblent ensuite se repousser, surtout au niveau des centromères, mais sont retenus ensemble par des zones appelées chiasmas, dans lesquelles les chromatides homologues sont attachées l'une à l'autre et où s'opèrent des échanges de matériel héréditaire entre chromatides homologues, phénomènes d'enjambement appelés aussi "crossing-over" (stade diplotène). Le résultat en est une recombinaison des gènes portés par chaque chromatide. En réalité, les échanges ont commencé lors de l'alignement des chromosomes homologues, mais les chiasmas ne deviennent visibles au microscope que lors de la phase de répulsion de ces chromosomes.
Exemple d'unique recombinaison génétique de chromatides homologues (rouge et verte) (à droite) consécutive à un enjambement (au milieu) entre deux chromosomes homologues (rouge et vert) rapprochés en métaphase de méiose 1 (à gauche).
En métaphase 1 les chromosomes migrent vers la plaque équatoriale. Contrairement à ce qui se passe au cours de la mitose, les centromères ne se dédoublent pas.
Lors de l'anaphase, les chromosomes homologues de chaque paire migrent de façon aléatoire vers un pôle cellulaire différent. Les deux cellules qui résultent de la télophase 1 sont donc haploïdes, puisqu'elles contiennent un unique jeu complet de chromosomes doubles, chaque chromosome étant d'origine paternelle ou maternelle. Il y a donc eu réduction de moitié du nombre de chromosomes par cellule. C'est pourquoi la première division de méiose est appelée division réductionnelle.
Chez certaines espèces, l'enveloppe nucléaire qui avait diparu en prophase 1 se reforme et la chromatine se déroule incomplètement. Dans ce cas a lieu une intercytokinèse, semblable à l'interphase de la mitose. Dans tous les cas, les chromatides constituant chacun des chromosomes double de la cellule ainsi formée ne sont plus semblables puisque des échanges génétiques ont eu lieu.

Différents stades de la méiose: seuls les chromosomes sont représentés. A=leptotène, B=zygotène, C=pachytène, D=diplotène, E=diacinèse, F=métaphase 1, G=anaphase 1, H=prophase 2, I=métaphase 2, J=anaphase 2, K=fin de télophase 2.
2.2 La méiose 2 ou division équationnelle
Pendant l'interphase qui précède la seconde division, il n'y a pas de duplication des chromosomes, car chacun d'eux s'était déjà dupliqué avant la méiose 1.
La méiose 2 est fort semblable à la mitose. Les chromosomes se disposent, au cours de la métaphase 2, dans la plaque équatoriale de chaque cellule haploïde. Les chromatides, chacune avec son centromère, se séparent et migrent vers les pôles cellulaires au cours de l'anaphase 2. Elles sont non identiques suite aux recombinaisons génétiques de la méiose 1.
Au terme de la télophase de la seconde division de méiose ou division équationnelle, on obtient quatre cellules-filles haploïdes possédant chacune un simple jeu de chromosomes non dupliqués.

Schémas illustrant les différents stades de la méiose d'une cellule végétale: la première ligne illustre l'interphase et la prophase de la division réductionnelle d'une cellule végétale (2n=4); la deuxième ligne illustre la métaphase et un enjambement chromosomique (1er dessin), l'anaphase (2è dessin) et la télophase (3è dessin) de la division réductionnelle, puis l'anaphase (4è dessin) et la télophase (5è dessin) de la division équationnelle. Les chromosomes homologues sont de couleurs différentes et les chromatides sœurs d'un chromosome double de même couleur. La troisième ligne illustre une autre possibilité de division réductionnelle (puis de division équationnelle): au cours de la méiose 1, le mélange des informations héréditaires est le fruit non seulement de la recombinaison chromosomique et mais aussi du mélange des chromosomes d'origines paternelle (en bleu) et maternelle (en rouge).
3 Comparaison entre la mitose et la méiose
Il convient de bien distinguer méiose et mitose:
|
Mitose |
Méiose |
|
Division équationnelle qui sépare les chromatides sœurs |
Une première division, réductionnelle, sépare les chromosomes homologues; les chromatides sœurs ne sont séparées qu'au cours d'une seconde division, équationnelle. |
|
Une division par cycle. |
Deux divisions par cycle. |
|
Pas d'appariement des chromosomes homologues; de ce fait, on n'observe ni enjambement, ni échange de matériel héréditaire (=recombinaison). |
Les chromosomes homologues s'apparient; on observe des enjambements; du matériel héréditaire peut être échangé entre chromatides homologues: c'est la recombinaison génétique. |
|
Une cellule-mère produit deux cellules-filles par cycle. |
Une cellule-mère produit quatre gamètes ou spores par cycle. |
|
Tous les produits portent la même information héréditaire. |
L'information héréditaire diffère d'un produit à l'autre, par mélange des chromosomes du parent mâle et des chromosomes du parent femelle, et par des recombinaisons génétiques. |
|
Les cellules-filles ont autant de chromosomes que la cellule-mère. |
Les cellules-filles ont moitié moins de chromosomes que la cellule-mère. |
|
Les cellules issues de la mitose sont en général capables de subir de nouvelles mitoses. |
Les cellules issues de la méiose ne peuvent subir une autre méiose, mais éventuellement bien des mitoses. |
|
La mitose se produit généralement dans presque toutes les cellules somatiques. |
La méiose ne se produit que dans les cellules spécialisées de la lignée germinale. |
|
Les mitoses commencent dès le stade du zygote et continuent tant que l'organisme est en vie. |
La méiose ne se produit qu'à maturité chez les organismes supérieurs (animaux, plantes à fleurs, conifères, ptéridophytes), mais elle apparaît dans le zygote de nombreuses algues et champignons. |
4 Avantage évolutif de la sexualité et de la méiose
Les phénomènes de reproduction et de sexualité ne sont pas systématiquement liés. La reproduction, seule présente dans les actes de reproduction asexuée, a pour seul objectif de multiplier les individus et de perpétuer l'espèce.
La sexualité est souvent associée à l'acte de reproduction (on parle alors de reproduction sexuée), mais elle a parfois lieu indépendamment, comme chez certains protozoaires et protophytes . Il s'agit dans ce cas d'un contact (=la conjugaison ) entre deux individus de types sexuels différents, au cours duquel les deux organismes s'échangent des copies de leur génome . Cet acte sexuel est une source de diversification du patrimoine héréditaire de l'individu, conférant à l'espèce une plus grande chance d'adaptation lors des modifications éventuelles des conditions du milieu de vie.
Chez les organismes supérieurs, la diversification du patrimoine génétique de l'espèce est assurée par l'union de deux gamètes, dont chacun porte un génome original à l'issue de la recombinaison génétique et de la répartition aléatoire des chromosomes maternels et paternels lors de la méiose précédant la formation des gamètes.
5 La gamétogenèse humaine
5.1 La spermatogenèse
La spermatogenèse a lieu durant toute la vie de l'homme dans les tubules séminifères où la maturation des cellules souches appelées spermatogonies commence à la puberté. La première division de méiose transforme chaque spermatocyte 1 diploïde en deux spermatocytes 2 haploïdes, la méiose 2 produisant à partir de ces deux cellules quatre spermatides, qui se transforment sans division en spermatozoïde.
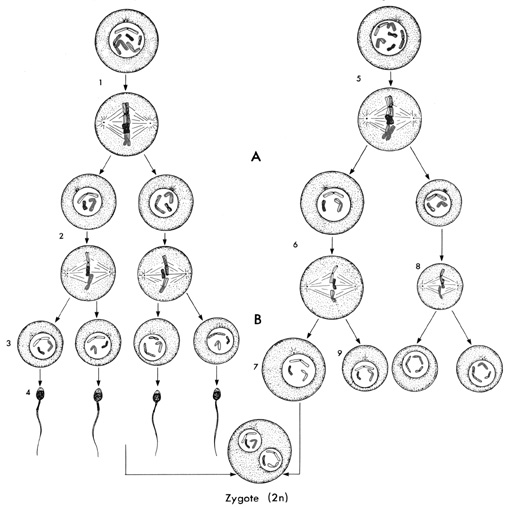
Spermatogenèse (à gauche) et o(v)ogenèse (à droite). A=division réductionnelle ou méiose 1. B=division équationnelle ou méiose 2. 1=spermatocyte 1. 2=spermatocytes 2. 3=spermatides (haploïdes). 4=spermatozoïdes. 5=O(v)ocyte 1. 6=O(v)ocytes 2. 7=O(v)otides. 8=premier globule polaire. 9=deuxièmes globules polaires.
5.2 L'ovogenèse
La diacinèse est une phase de repos inexistante ou très brève dans la spermatogenèse, mais de longue durée dans l'o(v)ogenèse, et comportant une réapparition de l'activité du nucléole, qui cessera à la métaphase 1. L'o(v)ocyte 1 de mammifère connaît en effet une longue pause entre le début de la première division de méiose, qui commence dans l'espèce humaine au troisième mois de la vie embryonnaire pour se bloquer en métaphase, et l'ovulation, qui ne peut avoir lieu qu'à partir de la puberté. Ce n'est qu'avec l'entrée d'un spermatozoïde dans l'ovule (en réalité au stade o(v)ocyte 2 haploïde) que prend place la seconde division de méiose.
Des quatre o(v)otides naissant des deux divisions de méiose d'un o(v)ocyte 1, trois sont petits et dégénèrent: ce sont les globules polaires. Le quatrième, beaucoup plus gros est l'ovule.
Contrairement à ce qui se passe chez l'homme, la multiplication des cellules souches que sont les o(v)ogonies est définitivement terminée alors que la petite fille est encore dans l'utérus maternel. Toutes les o(v)ogonies sont devenues des o(v)ocytes 1 à la naissance du bébé. La maturation des cellules germinales commence à la puberté et s'arrête à la ménopause.